Multiple independent appearances of the cecal appendix in mammalian evolution and an investigation of related ecological and anatomical factors.
Heather F. Smith, William Parker, Sanet H. Kotze, Michel Laurin.
Here is a link to the paper (pdf): https://drive.google.com/file/d/0B653CwHoRBNWXdtX0hWWDE3MDg/view?usp=sharing
Objectives
In this paper, scientists attempted to determine whether the presence of an appendix is correlated with other anatomical and ecological variables and to also obtain an accurate estimate of how many times the appendix evolved. The hypothesis was that the evolution of the appendix in animals was not associated with a decreasing cecum size as was previously assumed but instead associated with particular aspects of social behaviours or dietary factors which might increase the survival of advantageous intestinal bacteria.
Materials and Methods
The first step involved data compilation using a broad literature search of published studies on mammalian gastrointestinal anatomy. In order to correlate the presence of an appendix with ecological and anatomical variables, anatomical data was compiled at the species level. The anatomical data compiled were:
- appendix presence and length
- cecal size and morphology
- colon length
- presence of colonic separation mechanisms
- stomach histological lining (either entirely glandular or some squamous epithelium)
In order to correlate the presence of an appendix with dietary factors, data on the dietary categories of the species was compiled. In addition, data on the activity pattern, body mass and mean social group size for each species was obtained. Scientists also collected data on whether each species practiced coprophagia: the regular eating of any type of feces in sufficient amounts to contribute to the nutrition of the animal.
Scientists scored diet type into a binary character in which state 0 represents a cellulose-poor diet (including carnivory, insectivory, omnivory and frugivory) and state 1 represents cellulose-rich diet (including folivory, granivory and gummivory).
In order to identify the cecal appendix in animals, the cecal appendix was defined as a narrow, close-ended extension off the apex of the cecum with a distinct change in diameter of the lumen between it and the cecum. To determine whether the cecal appendix was more frequently found in combination with any particular cecal shape, the cecal morphology was characterized according to the following states: 0, cecum absent; 1, appendix-like; 2, spherical; 3, cylindrical; 4, tapering; 5, spiral; 6, paired.
To compare the relative sizes of the various parts of the digestive system in each of the 361 species, scientists divided linear measurements by the cubic root of body mass estimates they collected from literature.
A phylogenetic tree was constructed which includes 361 animal species representing all mammalian clades. The tree contained 19 polytomies (nodes) each with three to thirteen daughter branches.
Scientists in this study had both pairwise and continuous data therefore Pairwise Comparison tests were used to effectively evaluate the association between two changes in variables. All tests of correlations were made using pairwise comparison.
Scientists in the study also used the process of Parsimony Optimization: a procedure which minimizes the changes on a tree to account for character state distribution.
To compare the evolutionary rates of the appendix in various clades in the phylogenetic tree, scientists divided the number of inferred transitions (using parsimony) in each clade by that clade's sampled phylogenetic diversity index. The phylogenetic diversity index was calculated using the Stratigraphic Tools of Mesquite.
Results
Appendix Presence
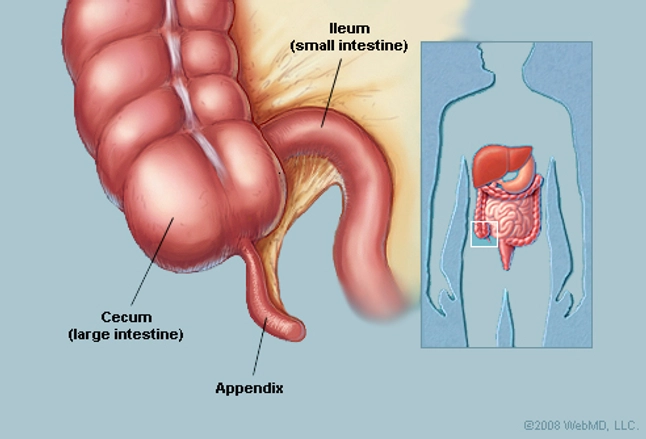
Of the 361 mammalian species sampled, 50 were found to have a cecal appendix. Several morphologies of the appendix were also found, see figure 1 for examples. Species in the clades Metatheria and Euarchontoglires were found to have an appendix. While species in the clade Laurasiatheria were found not to contain an appendix.

Source: H.F. Smith et al. / C. R. Palevol 12 (2013) 339–354.
Figure 1: The morphology of the cecal appendix in a variety of mammals. The black line indicates the approximate boundary of the appendix. (A) Human, Homo sapiens; (B) Orangutan, Pongo pygmaeus; (C) Southern hairy-nosed wombat, Lasiorhinus latofrons; (D) Echidna, Tachyglossus aculeatus; (E) Platypus, Ornithorhynchus anatinus; (F) Rabbit, Oryctolagus cuniculus; (G) Ground cuscus, Phalanger gymnotis; (H) Common brush-tail possum, Trichosurus vulpecula; (I) Cape dune mole-rat, Bathyergus suillus; (J) Brush-tailed porcupine, Atherurus africanus; (K) Beaver, Castor canadensis; (L) Koala, Phascolarctos cinereus; (M) Florida manatee, Trichechus manatus.
As seen in Figure 1, various morphologies of the cecal appendix were present:

As seen in Figure 1, various morphologies of the cecal appendix were present:
- Appendix-like as seen in C, D and E
- Spherical as seen in A, B and J
- Cylindrical as seen in I, K and M
- Tapering as seen in F, G, H and L
- Spiral and Paired morphologies were not pictured
Phylogenetic Signal
Table 1 shows the probability that the co-variation between character and phylogeny is random, this was assessed by comparing the number of steps (for discrete characters) or squared length (for continuous characters) over the reference tree to a population of 1000 random trees. Table 1 also shows the False Discovery Rate (FDR) threshold; the corrections for multiple testing were done through the False Discovery Rate. The sample size is the number of species in which the character is scored.
Source: H.F. Smith et al. / C. R. Palevol 12 (2013) 339–354.
Table 1: Phylogenetic Signal in the analyzed characters.
The results show that most characters analyzed displayed a strong phylogenetic signal that is the characters analyzed are strongly related to the phylogenies constructed. This confirms that phylogeny-based analysis such as pairwise comparisons are required to assess any correlations between the characters. The results also confirm that parsimony optimizations can reliably be used to infer character history.
The results showed that all three parts of the digestive system were positively correlated with each other. The correlation between the cecum and colon length was the strongest (P<0.001), whereas the correlation between the appendix and colon was the weakest (P=0.0170) although still significant. These correlations hold partly because most species without an appendix appear to have a smaller colon and cecum than species with an appendix. However, the scientists were not able to determine whether the correlation holds true within species with an appendix due to the small sample size.
The results of the pairwise comparison test revealed no statistically significant correlations between changes in the appendix and changes in colonic separation mechanisms, cellulose in the diet, stomach wall histological composition or cecal haustrations (small pouches which give the colon a segmented appearance) during the course of evolution.
The results also showed that highly social animals living in large groups and/or diurnal animals do not appear to be more likely to have an appendix compared to species living in small groups.

As seen in the results in Table 2, the appendix was found to have undergone 38 evolutionary events, including 32 to 38 gains and a maximum of six losses. The scientists interpreted the trend towards the appearance of an appendix as providing support to the hypothesis that the appendix is selectively advantageous to most mammals because if it were selectively neutral, losses should be about as common as gains.
The scientists concluded that the appendix is unlikely to serve any digestive function due to its small and narrow structure. Furthermore, it was difficult to support an other evolutionary scenario beyond its function as an immune structure.
In conclusion, the scientists concluded that the evolution of the appendix does not appear to be strictly tied to individual factors such as changes in diet, sociality or cecal-reduction across all clades. Although there were some caveats for this conclusion, such as:
Correlation between the evolution of the appendix and the evolution of other characters
Initially, Darwin suggested that the appearance of the hominoid appendix occurred simultaneously with the decrease in cecum size and a shift from folivory to frugivory, that is a decrease in cellulose consumption in the diet (Darwin, 1871).
The results from parsimony optimization confirms that the appearance of the appendix in hominoids is associated with a decrease in cecum size. However, no statistical test was performed to assess the probability that this association is random. These changes were also not associated with changes in the state of cellulose richness in the diet or changes in cellulose digestion.
In mammals, the only pattern consistent with Darwin's hypothesis occurs only in Trichechus manatus. Some catarrhine primates (especially hominoids) exhibit the predicted combination of an appendix accompanying a small cecum
Darwin's hypothesis predicts a reverse relationship between size of the appendix and cecum in mammals however scientists found a fairly strong positive correlation (P=0.005) between changes in the relative lengths of the appendix, cecum and colon in mammals.
Based on the results of parsimony optimization of appendix presence and cecum presence and size, the suggestion that appendix appearance is linked to a decrease in cecum size within mammals is refuted. Based on this, the scientists reconstructed the ancestral mammal and monotreme to have a small cecum.
Association between various characters and the evolution of the cecal appendix
The scientists tested the relationship between appendix and colon size, and between cecum and colon size in order to evaluate other relationships which might provide insight into the evolution of the morphology of the proximal colon.
The results showed that all three parts of the digestive system were positively correlated with each other. The correlation between the cecum and colon length was the strongest (P<0.001), whereas the correlation between the appendix and colon was the weakest (P=0.0170) although still significant. These correlations hold partly because most species without an appendix appear to have a smaller colon and cecum than species with an appendix. However, the scientists were not able to determine whether the correlation holds true within species with an appendix due to the small sample size.
The results of the pairwise comparison test revealed no statistically significant correlations between changes in the appendix and changes in colonic separation mechanisms, cellulose in the diet, stomach wall histological composition or cecal haustrations (small pouches which give the colon a segmented appearance) during the course of evolution.
The results also showed that highly social animals living in large groups and/or diurnal animals do not appear to be more likely to have an appendix compared to species living in small groups.
Rate of evolution of the cecal appendix

Source: H.F. Smith et al. / C. R. Palevol 12 (2013) 339–354.
Table 2: Evolutionary rates of the appendix in several major mammalian clades
As seen in the results in Table 2, the appendix was found to have undergone 38 evolutionary events, including 32 to 38 gains and a maximum of six losses. The scientists interpreted the trend towards the appearance of an appendix as providing support to the hypothesis that the appendix is selectively advantageous to most mammals because if it were selectively neutral, losses should be about as common as gains.
The scientists concluded that the appendix is unlikely to serve any digestive function due to its small and narrow structure. Furthermore, it was difficult to support an other evolutionary scenario beyond its function as an immune structure.
In conclusion, the scientists concluded that the evolution of the appendix does not appear to be strictly tied to individual factors such as changes in diet, sociality or cecal-reduction across all clades. Although there were some caveats for this conclusion, such as:
- The scientists used group size as a measure of sociality; however group size alone does not capture all aspects of sociality.
- The considerations of dietary intake do not take into account other important aspects of digestion such as the efficiency of energy extraction from ingested food.
Nevertheless, the scientists interpreted the trend towards the appearance of the appendix as providing support to the hypothesis that the appendix is selectively advantageous in most mammalian taxa because if it were selectively neutral, losses should be about as common as gains.
Critique
This paper by Smith et al. was very well structured. The introduction, materials and methods, results, discussion and conclusion were well formatted and easy to follow. All the figures were adequate for the results obtained. However, in the figures for the phylogeny trees shown in the paper, the species names were very hard to see which made it difficult to follow what was going on. Furthermore, the figures of the phylogeny trees were not thoroughly explained which made it even more difficult to understand the results obtained.
Most of the results from this paper were made using statistical analysis. Although this is a reliable means of interpreting data, it is difficult for readers without an advanced statistical knowledge to understand this paper.
In terms of the methods used in this experiment, the data used was from a compilation of a broad literature search of published studies. This led to some missing details as there was some data currently missing in the database. This led to a lack of power (the probability that the test correctly rejects the null hypothesis (H0) when the alternative hypothesis (H1) is true) in the statistical analysis. In particular, the lack of power was shown to affect a few of the tests, especially those concerning the concentration of lymphoid tissue in the cecum and appendix, colonic separation mechanism and coprophagia. The scientists however did note this and stated that the inclusion of additional species and additional data in the future could reveal patterns which were undetected by the present analysis.
Overall, the experiments seemed to be well conducted in this paper. However, there are a few things which the authors could have done differently. For example, in the results from parsimony optimization, no statistical test was performed to assess the probability that the association of the appearance of the appendix in hominoids along with a decrease in cecum size was random. In the future, perhaps the authors could statistically assess this probability. Additionally, the scientists were not able to determine whether the correlation between appendix and colon size, and between cecum and colon size holds true within animals in a species due to the small sample size. In the future, a larger sample size could be used.
Overall, the experiments seemed to be well conducted in this paper. However, there are a few things which the authors could have done differently. For example, in the results from parsimony optimization, no statistical test was performed to assess the probability that the association of the appearance of the appendix in hominoids along with a decrease in cecum size was random. In the future, perhaps the authors could statistically assess this probability. Additionally, the scientists were not able to determine whether the correlation between appendix and colon size, and between cecum and colon size holds true within animals in a species due to the small sample size. In the future, a larger sample size could be used.
The authors also refuted Darwin's hypothesis of the appendix however I do not think that the scientists can confidently refute this hypothesis. In this study, hominoids were found to possess an appendix associated with a small cecum, this pattern is consistent with Darwin's hypothesis. A similar pattern was not observed in other mammalian clades, this indicates that Darwin's hypothesis cannot be applied to other clades. However, it is important to note that Darwin formulated his hypothesis regarding the evolution of the appendix based on his observations in humans and other hominoids; therefore Darwin's hypothesis is partly correct as the results of this study seem to corroborate the first half of his hypothesis. It would have been more appropriate for the scientists to not completely refute his hypothesis but instead to modify it to be more inclusive of other mammalian clades.
In conclusion, although the results of this paper refute the hypothesis that the evolution of the appendix is associated with individual factors such as particular aspects of diet, social behaviours or dietary factors. In the future, perhaps scientists could test the possibility that some combinations of these could have a significant effect or test other factors which are equally as important.
In conclusion, although the results of this paper refute the hypothesis that the evolution of the appendix is associated with individual factors such as particular aspects of diet, social behaviours or dietary factors. In the future, perhaps scientists could test the possibility that some combinations of these could have a significant effect or test other factors which are equally as important.
References
Darwin, C. (1871). The descent of man and selection in relation to sex. John Murray, London.
Smith, H., Parker, W., Kotzé, S. H., & Laurin, M. (2013). Multiple independent appearances of the cecal appendix in mammalian evolution and an investigation of related ecological and anatomical factors. Comptes Rendus - Palevol, 12(6): 339-354.
Smith, H., Parker, W., Kotzé, S. H., & Laurin, M. (2013). Multiple independent appearances of the cecal appendix in mammalian evolution and an investigation of related ecological and anatomical factors. Comptes Rendus - Palevol, 12(6): 339-354.